
La proliferación de aguas residuales desencadena la floración de Trichodesmium y aumenta las tasas de fijación del Nitrógeno

En SAN BORONDÓN: El presente artículo ha sido traducido al español desde su fuente original, la prestigiosa revista NATURE, en https://www.nature.com/articles/s41598-017-04622-8 . El interés de este documento es que contradice frontalmente los argumentos utilizados por el Gobierno de Canarias, especialmente los publicados en su web www.canariassaludable.org, que pretenden desvincular de forma interesada los vertidos ilegales de aguas fecales sin tratar en nuestras costas con los blooms de cyanobacterias.
La proliferación de aguas residuales desencadena la floración de Trichodesmium y aumenta las tasas de fijación del Nitrógeno
El presente artículo ha sido traducido al español desde su fuente original, la prestigiosa revista NATURE, en https://www.nature.com/articles/s41598-017-04622-8 . El interés de este documento es que contradice frontalmente los argumentos utilizados por el Gobierno de Canarias, especialmente los publicados en su web www.canariassaludable.org, que pretenden desvincular de forma interesada los vertidos ilegales de aguas fecales sin tratar en nuestras costas con los blooms de cyanobacterias.
NOTA DEL TRADUCTOR: Para evitar confusiones entre la nomenclatura y símbolos de la formulación química y las referencias a la bibliografía, toda vez que el documento original se utiliza la notación superindices para ambos casos, cuando se produjo ambigüedad en el contexto de la traducción se encerró entre corchetes “[] “ las referencias a la bibliografía.
La proliferación de aguas residuales desencadena la floración de Trichodesmium y aumenta las tasas de fijación de N2
Resumen:
El Mediterráneo meridional (SEMS) es un ambiente marino cálido y soleado con baja concentración de N ambiente, por lo que se considera ideal para la diazotrofía por diazotrófos autotróficos como Trichodesmium. A pesar de las condiciones favorables, las tasas de fijación de N2 son a menudo bajas y apenas ha sido detectado Trichodesmium en el SEMS. Este estudio informa sobre la aparición de una floración de Trichodesmium en el SEMS que se atribuyó a T. erythraeum de acuerdo con la huella dactilar de ADN del gen nifH. Se encontró que esta floración (1407 ± 983 células L-1) fue desencadenada por una intensa explosión de aguas residuales crudas que suministraron altas concentraciones de N, P y carbono orgánico disuelto (DOC), lo que resultó en una baja N: P (~ 12: 1) y relaciones excepcionalmente altas C: P (~ 1340: 1). Supusimos que estas condiciones proporcionaron condiciones favorables para que la floración de Trichodesmium se formase a través del metabolismo mixotrófico. Como resultado, se registró un aumento de cuatro veces en la fijación de N2, que contribuyó con ~ 70% a la nueva producción primaria y estimuló un fuerte aumento en la actividad del fitoplancton y la biomasa. Las conclusiones de este estudio apuntan a un nuevo paradigma para la formación de cyanobacterias T. erythraeum que está estrechamente vinculado a fuentes antropogénicas y pronta productividad microbiana en ambientes marinos oligotróficos como el SEMS.
Introducción
Estas escorrentías abruptas a menudo no son tratadas y contienen patógenos, nutrientes orgánicos e inorgánicos, metales pesados y detergentes, por lo que se consideran amenazas importantes para los ecosistemas marinos3, Por ejemplo, los estallidos de aguas residuales introducen nutrientes que pueden estimular los efectos de abajo hacia arriba7 y desplazar la estructura de la comunidad del fitoplancton para formar grandes floraciones de dinoflagelados tóxicos u otros Especies de algas dañinas8,9,10.
El agua superficial del Mediterráneo meridional (SEMS) es un ambiente marino cálido y soleado con condiciones ultra-oligotróficas, que resulta en baja biomasa bacteriana y fitoplanctónica y baja productividad11,12,13. La productividad del fitoplancton se considera N-limitada14 o N-P-co-limitada15,16, mientras que las bacterias son P-limitadas17 o C-limitadas14. Estas condiciones son supuestamente ideales para que la diazotrofia se produzca por diazotrofos autotróficos que pueden fijar C y N usando la energía de la luz solar18. Aunque la comunidad diazotrófica en el Mediterráneo oriental es muy diversa14,19,20, las tasas típicas de fijación de N2 son constantemente bajas (<0,4 nmol NL-1 d-1) 13,14,21,22,23, mientras que la Trichodesmium spp sólo se notificaron una vez en el Mediterráneo noreste24 y recientemente en el Golfo de Gabes (Túnez) 25.
Trichodesmium spp.
Se considera que es uno de los microorganismos dominantes de fijación de nitrógeno en sistemas marinos26,27. Se informó previamente que las grandes floraciones de Trichodesmium pueden fijar CO2 y N2 a altas velocidades alcanzando 640 pg de células C 1 d-1 y 29 pg de células N -1 d-1, respectivamente, induciendo así la banda alimentaria microbiana 28. La formación de bloques de Trichodesmium a menudo se ve obstaculizada por la disponibilidad de metales traza (principalmente Fe) y otros nutrientes (principalmente P) 29, 30, temperatura del agua31, disponibilidad de luz32, 33, altos niveles de O2[34], así como otras características fisicoquímicas del agua35. Sin embargo, las condiciones que restringen la formación de floración enel SEMS de Trichodesmium spp., que es potencialmente ideal para diazotrophy, son actualmente desconocidas 18.
En este estudio, nos centramos en los posibles vínculos entre el arranque de aguas residuales y el desarrollo de blooms de Trichodesmium y la correspondiente estructura de la comunidad microbiana. Con este fin, seguimos una floración de Trichodesmium erythraeum en el agua costera de la SEMS que fue desencadenada por una explosión de aguas residuales municipales. Durante este evento, seguimos la dinámica temporal de T. erythraeum y la fijación de N2, así como picofitoplancton, diatomeas, dinoflagelados y bacterias heterotróficas. Nuestros resultados apuntan a los posibles impactos de las corrientes de aguas residuales que fluyen en ambientes marinos ultra-oligotróficos como el SEMS.
Materiales y métodos.
Lugar de estudio y colección de muestras.
Durante el invierno se recolectaron muestras de agua de mar (~ 20 L) de las aguas superficiales (~ 0,5 m) de la costa sureste del Mediterráneo (32 ° 49'34 N, 34 ° 57'20 E). Se realizaron hasta diez mediciones; (ii) cinco durante el evento, y (iii) dos inmediatamente después de que el estallido de aguas residuales hubiera terminado. El agua de mar fue analizada buscando nutrientes inorgánicos, carbono orgánico disuelto, clorofila-a (como un proxy de algas), abundancia de picofitoplancton incluyendo Synechococcus + Prochlorococcus (colectivamente denominados cianobacterias autotróficas) y picoeucariotas, abundancia de bacterias heterotróficas y abundancia de microphytoplankton (diatomeas y dinoflagelados). Además, se recogieron muestras de agua para Trichodesmium spp. Abundancia, diversidad diazotrófica (análisis de genes nifH), producción primaria, producción bacteriana y medidas de fijación de N2.
Los nutrientes inorgánicos y el carbono orgánico disuelto (DOC)
Se recogieron muestras de agua para las concentraciones de NO2 + NO3, PO4 y Si (OH)4 en 15 ml de viales de centelleo de plástico lavados con ácido y se colocaron inmediatamente en un congelador a -20ºC hasta su análisis. Los valores de nutrientes inorgánicos se determinaron utilizando el sistema segmentado Seal Analytical AA-3[36].
Las muestras para las concentraciones de DOC (40 ml) se recogieron en viales de vidrio con tapón septum y se acidificaron con ácido clorhídrico concentrado (32%) en una proporción de 1: 1000 y se almacenaron en la oscuridad a 4º C hasta su análisis. Las muestras se analizaron en un analizador Shimadzu TOCV con una precisión de 2 μmol L-1.
Clorofila - a (Chl – a)
Se filtraron muestras de agua de mar (300 mL) a través de un filtro Whatman GF / F (~ 0,7 \ μm de tamaño de poro nominal) y se mantuvieron a -20ºC en la oscuridad. Los filtros se extrajeron durante la noche en acetona (90%) y se determinaron por el método de no acidificación37 usando un fluorómetro de Turner Designs (Trilogy) con filtros de emisión de excitación de 436-nm y de 680-nm.
Concentración de Picophytoplankton
Se fijaron muestras de agua (1,7 ml) con glutaraldehído al 50% (Sigma-Aldrich G7651), se congelaron instantáneamente en nitrógeno líquido y se almacenaron a -80ºC. La concentración de picofitoplancton se determinó en base a la auto-floración naranja de la ficoeritrina y la auto-floración roja del chl-a38. La abundancia bacteriana heterotrófica se midió teñiendo la muestra con 1 μl de SYBR verde (Applied Biosystems cat # S32717) seguido de una incubación de 10 minutos en la oscuridad. Todas las muestras se analizaron mediante un citómetro de flujo de enfoque acústico Attune® (Applied Biosystems) equipado con láseres de 488 nm y 405 nm. Para el tamaño estándar, se utilizaron perlas de 1 μm (Polysciences).
Abundancia de trichodesmio, diatomeas y dinoflagelados
Las densidades de células se determinaron después de concentrar 1-5 l de agua de mar superficial y contar tres submuestras. Los números de células se estimaron utilizando una Sedgewick-Rafter Cell (S50) y un microscopio de fluorescencia de luz (Olympus BH-2) con una ampliación de 20-40X.
Extracción y secuenciación del gen nifH
Las muestras (1-5 L) se filtraron a través de 0,2 micras filtros Supor (PALL Corp.) y se colocaron en una bolsa estéril DNasa / RNasa libre Whirl-Pak. Las muestras se congelaron instantáneamente en nitrógeno líquido y se almacenaron a -80ºC. El ADN se extrajo utilizando el método fenol-cloroformo según Man-Aharonovich19. Nitrogenase Fe transcripciones de proteínas (nifH) se amplificaron mediante una estrategia de anidación PCR39. Se realizó una secuenciación en pares de ADN de ADN en una plataforma Illumina MiSeq en los Laboratorios de Investigación y Ensayo (Lubbock, TX, EE.UU.).
Análisis secuencial
Las lecturas de Illumina fusionadas se filtraron y se analizaron con el pipeline de información cuantitativa en ecología microbiana (QIIME )40. Las lecturas restantes se almacenaron en unidades taxonómicas operativas (OTU) y se definieron con un 97% de similitud utilizando el algoritmo UCLUST41. La taxonomía se asignó con BLAST y una base de datos de secuencias nifH de Heller et al.42. Los árboles filogenéticos se generaron con FastTree en QIIME43 y se visualizaron con los paquetes Interactive Tree of Life (IToL) y Topiary Explorer v1.0.
Producción primaria (PP)
Las tasas de fijación fotosintética de carbono se estimaron utilizando la incorporación de Carbono-14[44]. Las muestras de agua se analizaron por triplicado con controles oscuros y de tiempo cero. Se añadieron muestras (50 ml) recogidas en cada punto de tiempo a botellas de policarbonato (Nalgene) que contenían 5 μCi de NaH14CO3 (Perkin Elmer, actividad específica 56 mCi mmol-1) y se incubaron durante 4 h bajo iluminación natural y temperatura ambiente. Para determinar la cantidad de la radiactividad añadida, se mezclaron inmediatamente 50 μl de cada muestra con 50 μl de etanolamina y se almacenaron para análisis. Las incubaciones se terminaron filtrando el agua de mar con punta a través de filtros GF / F (~ 0,7 μm de tamaño de poro nominal) a baja presión (~ 50 mmHg). Los filtros se incubaron durante la noche en viales de centelleo de 5 ml que contenían 50 μl de HCl al 32% con el fin de eliminar el exceso de bicarbonato de Carbono-14. Después de añadir 5 ml de cóctel de centelleo (Ultima-Gold) a cada vial, se midió la radiactividad usando un contador de centelleo líquido TRI-CARB 2100 TR (Packard).
Producción bacteriana (BP)
Las tasas se estimaron utilizando el método de incorporación [4,5-3H] -leucina45. Se incubaron tres alícuotas (1,7 ml cada una) de cada muestra de agua con 100 nmol de leucina L-1 (Amersham, actividad específica 160 Ci mmol-1) durante 4 horas a temperatura ambiente en la oscuridad. Se utilizaron muestras como muestras de muestras tratadas con ácido tricloroacético (TCA). Las incubaciones se terminaron con 100 μl de TCA (100%), seguido por micro-centrifugación. Después de añadir 1 ml de cóctel de centelleo (Ultima-Gold) a cada vial, se contaron las muestras usando un contador de centelleo líquido TRI-CARB 2100 TR (Packard). Se utilizó un factor de conversión de 3 kg C por mol de leucina incorporada, suponiendo una dilución isotópica de 2.046.
Fijación del Dinitrógeno (N2)
Las tasas se midieron por triplicado usando el protocolo de agua de mar enriquecido47 con 15N2. El agua de mar enriquecida con 15N2 se preparó inyectando gas de 15N2 (99%) 1: 100 (vol: vol) en agua de mar desgaseada (MiniMódulo G543) y filtrada (Policarbonato 0,2 μm) recogida en el sitio de estudio. El stock de agua de mar enriquecido se agitó vigorosamente con el fin de disolver completamente el gas 15N2, y alícuotas (225 ml) se añadieron entonces a triplicar botellas experimentales de Nalgene (4,6 l). Después de 24 h de incubaciones bajo condiciones de luz y temperatura ambiente, las muestras se filtraron a través de filtros GF / F previamente quemados (450ºC, 4,5 h) y se secaron en un horno a 60ºC durante la noche. Las muestras se analizaron a continuación utilizando un analizador elemental CE Instruments NC2500 conectado a un espectrómetro de masas de proporción de isótopos Delta Plus XP de Thermo-Finningan (IRMS). Para la espectrometría de masas de relación de isótopos, se realizó una curva estándar para determinar la masa de N con cada corrida de muestra.
Análisis estadístico
Los datos se muestran como promedios; Las barras de error significan una desviación estándar (n = 3-5). Las relaciones entre las diferentes variables ambientales y fisiológicas se determinaron con una prueba de correlación de Pearson (P <0,05). Todas las pruebas se realizaron utilizando el software XLSTAT.
Resultados y discusión
Las corrientes de aguas residuales no tratadas en el medio costero del SEMS ocurren unas pocas veces cada invierno, generalmente debido a deficiencias en la red de alcantarillado y/o sobrecarga del sistema de drenaje de aguas residuales por las escorrentías de aguas pluviales. La introducción de cargas elevadas de materia orgánica y nutrientes en ambientes costeros oligotróficos como el SEMS a menudo puede dar lugar a la floración de fitoplancton (Fig. 1B, C y 2). La formación de estas floraciones puede alterar la dinámica microbiana de la red alimentaria48 y liberar diferentes toxinas dañinas49, así como obstaculizar el desempeño de la desalinización a gran escala y las centrales eléctricas50.
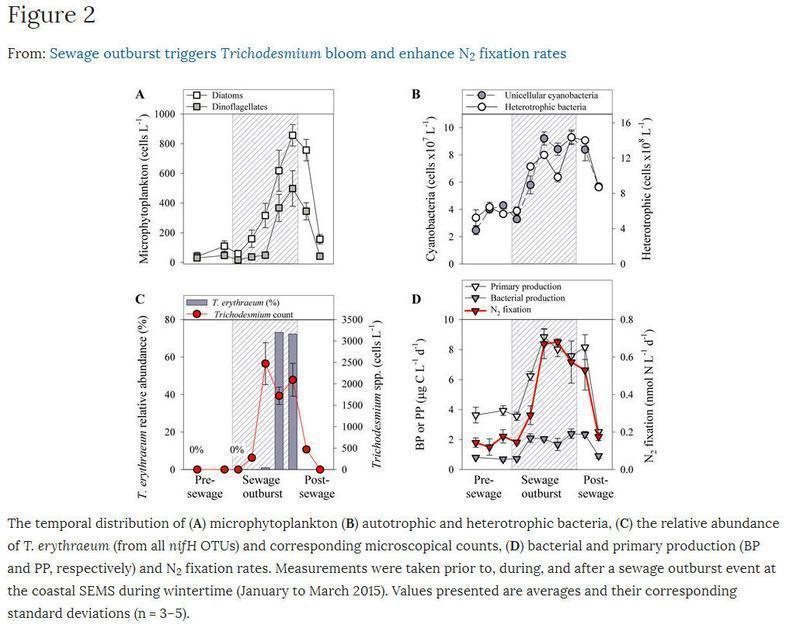
Condiciones ambientales del sitio de estudio durante el invierno
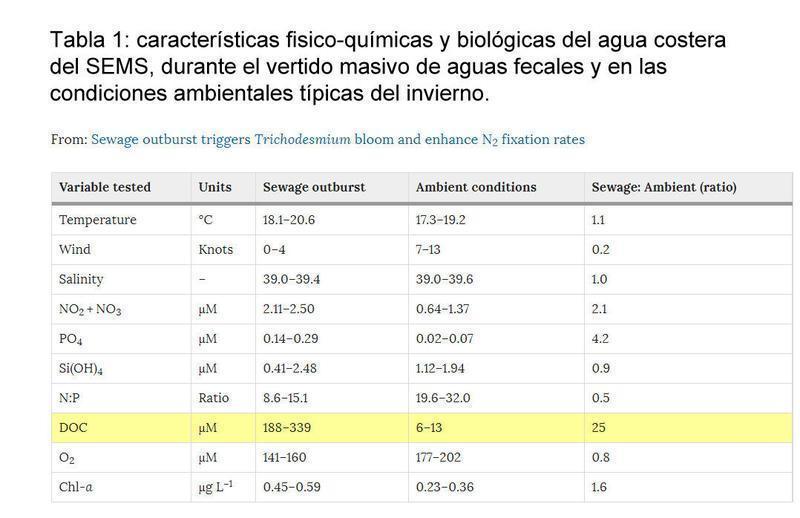
El agua costera antes del evento de desprendimiento exhibió características SEMS típicas de invierno (Tabla 1), con agua fría (18,3 ± 0,7 ° C), barrida por el viento (10 ± 3 nudos) y bien oxidada (190 ± 10 μM). Los niveles de nutrientes inorgánicos, es decir, NO2 + NO3, PO4 y Si (OH)4, fueron bajos (cercanos al límite de detección para P y hasta ~ 2 μM) así como de carbono orgánico disuelto (DOC, 10 ± 5 μM). Estas condiciones fisicoquímicas dieron lugar a una baja biomasa de fitoplancton (0,32 ± 0,05 μg chl-a L-1), que es típica para esta región14, 23. Synechococcus fue el fototropo más abundante (3,9 ± 1,2 células × 107 L-1), mientras que la diatomea (116 ± 43 células L-1, principalmente Chaetoceros spp.) Y dinoflagelados (34 ± 13 células L-1, en su mayoría especies de Ceratium) fueron 5-6 órdenes de magnitud más bajas (Fig. 2A, B). En este momento, Trichodesmium spp. no se pudo detectar (figura 2C). La baja abundancia fototrófica resultó en escasas tasas de PP (3,3 ± 0,6 μg C L-1 d-1, Fig. 2D). La abundancia de bacterias heterotróficas fue mayor de diez veces con respecto a Synechococcus (64 ± 14 células × 107 L-1, Figura 2B). Sin embargo, las tasas de PA fueron bajas (0,8 ± 0,2 μg C L-1 d-1) y constituyeron ~ 25% del PP (Fig. 2D). Las tasas de fijación de N2también fueron bajas (0,15 ± 0,02 nmol NL-1 d-1, Fig. 2D) y similares a los valores reportados previamente en este área de estudio14, 23. Atribuimos las bajas tasas de fijación de N2 a alfa, gamma y delta-proteobacteria Diazotróficos como se informó recientemente para la costera SEMS14.
Condiciones costeras en el SEMS durante un evento de desbordamiento de aguas residuales
Durante febrero de 2015, las aguas residuales crudas y no tratadas fueron continuas e intensas (miles de L h-1) descargadas durante cinco días en el agua costera de la SEMS debido a diversas alteraciones en el sistema municipal de drenaje de Haifa, Israel (Figura 1A) . Durante este período, la temperatura del agua y la salinidad no se vieron afectadas y se mantuvieron típicas de la estación, y sólo se registró una brisa débil (<4 nudos) durante el episodio de desagüe de aguas residuales (Tabla 1). Al mismo tiempo, los niveles de NO2 + NO3 (2,3 ± 0,2 μM) y PO4 (0,2 ± 0,1 μM) aumentaron en dos y cuatro veces respectivamente (Tabla 1). Este cambio en NO2 + NO3 y PO4 resultó en una reducción de la relación N: P (~ 12), que es menor que la proporción colonial de 16: 1 Redfield. Los niveles de DOC también se han incrementado en 25 veces con respecto a las condiciones ambientales pre-alcantarillado (268 ± 56 μM).
Después de la explosión de aguas residuales, una gran floración Tuff-like de Trichodesmium spp. Se desarrolló, alcanzando densidades de 1407 ± 983 tricomas L-1 (Figs 1B y 2C). Las huellas genéticas de nifH indicaron que la especie diazotrófica más abundante fue Trichodesmium erythraeum (T. erythraeum), constituyendo más del 70% de todas las OTU diazotróficas en la pluma de aguas residuales (Figura 2C). Al mismo tiempo, las OTU relativas de otros diazótrofos, tales como proteobacterias heterotróficas (es decir, Desulfobacter sp.) Y cianobacterias (es decir, Leptolyngbya sp.), eran significativamente más bajas (<1%). Nuestras densidades medidas de T. erythraeum estuvieron de acuerdo con las observadas anteriormente en el Mediterráneo noreste (Mar Egeo) en septiembre de 201024 y en una prospección de tres años en el Golfo de Gabes (Túnez) 25. Sin embargo, estos números de células estaban por debajo de las densidades reportadas para Trichodesmium spp a gran escala. Floraciones en otros ambientes marinos que oscilaron entre 3000 y 10000 tricomas L-1 51,52,53.
Junto con el fuerte y significativo aumento de T. erythraeum, las tasas de fijación de N2 aumentaron en ~ cuatro veces (Fig. 2D). Por lo tanto, atribuimos la parte del león de las tasas de fijación N2 correspondientes a T. erythraeum (Tabla 2). Esta relación produjo tasas de fijación de N2 específicas por tricoma de ~ 3,5 nmol N tricome-1 d-1, que se encuentran en el extremo inferior de los valores reportados de otros sistemas oceánicos donde Trichodesmium spp. Predomina53,54,55, o de los monocultivos de T. erythraeum56. Sin embargo, debe tenerse en cuenta que no todas las células dentro de un filamento de Trichodesmium tomar un papel activo en la fijación de N2 [34]. Por lo tanto, es posible que los filamentos de T. erythraeum observados aquí tuvieran menos células fijadoras de N2 (fuera de todas las células vegetativas) que en otros sistemas marinos, lo que resultaría en menores tasas de fijación específica de N2 en comparación con otros sistemas marinos.
Concomitantemente con el evento de desprendimiento de aguas residuales y la floración de T. erythraeum, los niveles de chl-a aumentaron positiva y linealmente en un 60% (0.51 ± 0.06 μg L-1) y PP en 230% (7.77 ± 0.97 μg CL-1 d-1) En comparación con las concentraciones ambientales (Tablas 1 y 2), lo que indica un aumento significativo de la biomasa fototrófica. Específicamente, estos cambios fueron aún más evidentes en el aumento de la abundancia de diatomeas (cuatro veces mayor de Asterionellopsis glacialis y Leptocylindrus danicus) y dinoflagelados (~ siete veces, mayor Ceratium spp.) tras el desarrollo de la floración de T. erythraeum (Figura 2A- C, Tabla 2). En contraste con el aumento significativo en la abundancia de microphytoplankton, bacterias heterotróficas sólo aumentó en 1,9 veces (Fig. 2B]; Sin embargo, las tasas de PA aumentaron tres veces con relación a las tasas de producción antes del alta. Este desacoplamiento entre el aumento moderado de las bacterias heterotróficas totales y el aumento significativo de las tasas de PA, junto con los altos niveles de DOC que se introdujeron con las aguas residuales (Tabla 1), sugiere que algunos fotótrofos también podrían utilizar estos sustratos orgánicos cambiando a la nutrición mixotrófica57 , 58.
La importancia de Trichodesmium para la nueva producción y la biomasa microbiana en el SEMS
Se sabe que las floraciones de trichodesmio liberan nitrógeno biodisponible en el agua circundante, dando lugar a un crecimiento mayor de cianobacterias no diazotróficas y microphytoplankton22. De hecho, la aparición de T. erythraeum en el agua SEMS se asoció positiva y significativamente con la biomasa fototrófica (como chl-a), la abundancia de Synechococcus, la fijación de PP y N2(Tabla 2). También se correlacionó positivamente con la abundancia de microphytoplankton, aunque no significativamente (Tabla 2]. Estimamos que la contribución de T. erythraeum a PP vía fijación de N2 (basada en la estequiometría de Redfield de C: N) alcanzó 68 ± 27%. Esta contribución es siete veces más alta que el porcentaje habitualmente notificado en la misma zona de estudio y temporada durante las condiciones de no floración (~ 10%) 23 y 30-140 veces mayor que el reportado de las aguas costa afuera del Mediterráneo oriental ( Sugerimos que el nuevo N derivado de la fijación de T. erythraeum N2, junto con los nutrientes adicionales que fueron introducidos por las aguas residuales, puede inducir tasas de producción de bacterias primarias y bacterianas, Lo que resulta en el crecimiento de microphytoplankton (Fig. 2, Tabla 2]. La abundancia de diatomeas y dinoflagelados aumentó notablemente al cuarto día de la explosión de las aguas residuales y a los 1-2 días desde la aparición de Trichodesmium (Fig. 2). Por lo tanto, supusimos que es probable que el crecimiento del microphytoplankton se ligó a Trichodesmium y las correspondientes tasas de fijación de N2 elevados. Del mismo modo, una floración Trichodesmium en las aguas cercanas a la costa de Goa (oeste de la India) llevó a Asterionella japonica y Nitzschia closterium blooms61. En el Golfo de México, Trichodesmium spp. Desencadenó una floración del dinoflagelado tóxico Karenia brevis62. Adicionalmente, los análisis recientes de nanoescala de espectrometría de masas de iones secundarios (NanoSIMS) mostraron que el picofitoplancton autotrófico de tamaño pequeño acumulaba Trichodesmium spp. En las aguas del suroeste del Océano Pacífico alrededor de Nueva Caledonia22. Además, se observó que las floraciones a gran escala de Trichodesmium disminuían rápidamente a través del pastoreo por los copépodos harpacticoides63, por lisis viral64 o por una muerte celular programada genéticamente controlada28, 53, 65. Estos desprendimientos de floración rápida (~ 50% de la pérdida de biomasa en 24 h) Pueden alimentar niveles tróficos más altos y cambiar significativamente la estructura de la comunidad microbiana.

Factores que controlan la actividad y la formación de bloom de Trichodesmium en el SEMS
Los factores ambientales, como la temperatura de la superficie del mar, la pCO2, la irradiancia, el estado de mar en reposo y la disponibilidad de nutrientes tales como P Y Fe, se han sugerido como factores de control para las floraciones de Trichodesmium29, 30, 33, 66, 67, 68, 68, 70. Actualmente, las razones por las que las floraciones de Trichodesmium no se desarrollan en el SEMS, a pesar de ser un entorno marino potencialmente ideal con características beneficiosas para la diazotrofia, todavía se desconocen. Nuestros hallazgos sugieren que aparte del mar en calma, las condiciones de luz solar y la relación N: P (12: 1) algo baja, dos mecanismos pudieron estar en juego y desencadenado el desarrollo de la floración de T. erythraeum: (i) Las altas concentraciones De N y P (a una baja relación N: P) que se introdujeron en el medio costero con las aguas residuales suministró las necesidades metabólicas celulares iniciales para el crecimiento y proliferación de células de T. erythraeum. Ii) La alta concentración de DOC que se suministró con las aguas residuales puede haber proporcionado una fuente de carbono asimilable al medio ambiente costero. Un estudio reciente mostró que Trichodesmium, como otros diazotróficos unicelulares71, 72, puede utilizar DOC como una fuente de carbono disponible73 y por lo tanto puede proporcionar una fuente de energía alternativa para fijar N2 y formar floraciones. Suponiendo que las células de T. erythraeum son mixotróficas73, sugerimos que las floraciones de Trichodesmium podrían ser estimuladas por este tipo de brotes de aguas residuales en el SEMS oligotrófico. Por otra parte, durante la floración, las altas concentraciones de DOC resultaron en una proporción excepcionalmente alta de C: N: P (~ 1340: 12: 1) con relación a la relación ambiental medida durante el período de no floración (~ 233: 25: 1) . Sugerimos que estas condiciones pueden haber inducido N-limitante todavía C-ricos condiciones que priorizan el metabolismo mixotrófico por Trichodesmium. Cabe señalar que, si bien Benavides et al.73 sugirieron que las poblaciones naturales de Trichodesmium pueden utilizar la nutrición mixotrófica alternativa y asimilar el DOC como una fuente primaria de C, tal adición no necesariamente desencadenaría una floración de Trichodesmium en el SEMS. De hecho, un estudio reciente de la SEMS costera informó que la diazotrofía heterotrófica fue estimulada por una adición de DOC + N + P, mientras que Trichodesmium spp. No se observó14. Proponemos que materiales adicionales como metales traza y vitaminas se introdujeron con el arranque de aguas residuales y desempeñó papeles mutuos en la formación de la floración de T. erythraeum que se informó aquí. Sin embargo, las razones de por qué floraciones de Trichodesmium spp. no se desarrollan más a menudo en el SEMS y el Mar Mediterráneo (a pesar de estar rutinariamente expuestos a los nutrientes orgánicos e inorgánicos externos) sigue siendo desconocido y merece más investigación.
Conclusiones
Nuestros resultados proporcionan la primera observación registrada de una floración de Trichodesmium (T. erythraeum) en las aguas costeras de la SEMS, que fue estimulada por un evento prolongado e intenso de desagüe de aguas residuales. Sugerimos que la introducción de altas cantidades de N, P y DOC, así como la relación C: N: P medida aquí, desencadenó la formación de la floración de Trichodesmium. Específicamente, suponiendo que Trichodesmium son mixotrofos, las altas concentraciones de DOC pueden haber proporcionado las fuentes de energía para cumplir con los requisitos metabólicos para la fijación de N2 y la formación de floración. Cabe señalar que una investigación más profunda y dedicada es el mérito para probar las estrategias de metabolismo de Trichodesmium.
Siguiendo lo anterior, nuestros resultados indican que la contaminación antropogénica, como las corrientes de aguas residuales sin procesar en entornos oligotróficos, puede no sólo desencadenar floraciones de T. erythraeum, sino también cambiar la estructura de la comunidad microbiana. Es probable que estos eventos tras grandes emisiones de aguas residuales puedan tener grandes implicaciones ecológicas a través de las floraciones de Trichodesmium en entornos oligotróficos como el SEMS. Sin embargo, insistimos en que se necesitan más experimentos controlados y monitoreo in situ para entender mejor la dinámica, la regulación y la formación de blooms de Trichodesmium en el SEMS.
Información Adicional
Nota del editor: Springer Nature permanece neutral con respecto a las reclamaciones jurisdiccionales en los mapas publicados y en las afiliaciones institucionales.
Referencias
Link para descargar las referencias en formato RIS clic aquí
1. Kirby, R. R. & Beaugrand, G. Trophic amplification of climate warming. Proc. Biol. Sci. 276, 4095–4103 (2009).
2. Yuan, Z., Shi, J., Wu, H., Zhang, L. & Bi, J. Understanding the anthropogenic phosphorus pathway with substance flow analysis at the city level. J. Environ. Manage. 92, 2021–2028 (2011).
3. Nixon, S. W. Coastal marine eutrophication: A definition, social causes, and future concerns. Ophelia 41, 199–219 (1995).
4. Halpern, B. S. et al. A global map of human impact on marine ecosystems. Science. 319, 948–952 (2008).
5. Hansson, S. et al. The stable nitrogen isotope ratio as a marker of food-web interactions and fish migration. Ecology 78, 2249–2257 (1997).
6. Rombouts, I. et al. Food web indicators under the Marine Strategy Framework Directive: From complexity to simplicity? Ecol. Indic.29, 246–254 (2013).
7. Davis, J. M., Rosemond, A. D., Eggert, S. L., Cross, W. F. & Wallace, J. B. Long-term nutrient enrichment decouples predator and prey production. Proc. Natl. Acad. Sci. USA 107, 121–126 (2010).
8. Michael Beman, J. et al. Agricultural runoff fuels large phytoplankton blooms in vulnerable areas of the ocean. Nature434, 211–214 (2005).
9. Anderson, D. M. et al. Harmful algal blooms and eutrophication: Examining linkages from selected coastal regions of the United States. Harmful Algae 8, 39–53 (2008).
10. Bauman, A. G., Burt, Ja, Feary, Da, Marquis, E. & Usseglio, P. Tropical harmful algal blooms: An emerging threat to coral reef communities? Mar. Pollut. Bull. 60, 2117–2122 (2010).
11. Berman, T., Townsend, D. & Elsayed, S. Optical transparency, chlorophyll and primary productivity in the eastern Mediterranean near the Israeli coast. Oceanol. Acta 7, 367–372 (1984).
12. Pulido-Villena, E., Wagener, T. & Guieu, C. Bacterial response to dust pulses in the western Mediterranean: Implications for carbon cycling in the oligotropic ocean. Global Biogeochem. Cycles22, 1–12 (2008).
13. Rahav, E. et al. Uncoupling between dinitrogen fixation and primary productivity in the eastern Mediterranean Sea. J. Geophys. Res. Biogeosciences 118, 195–202 (2013).
14. Rahav, E., Giannetto, M. & Bar-Zeev, E. Contribution of mono and polysaccharides to heterotrophic N2 fixation at the eastern Mediterranean coastline. Sceintific Reports 6, 27858 (2016).
15. Kress, N. et al. Effect of P and N addition to oligotrophic Eastern Mediterranean waters influenced by near-shore waters: A microcosm experiment. Deep. Res. Part II Top. Stud. Oceanogr. 52, 3054–3073 (2005).
16. Zohary, T. et al. P-limited bacteria but N and P co-limited phytoplankton in the Eastern Mediterranean—a microcosm experiment. Deep Sea Res. Part II Top. Stud. Oceanogr. 52, 3011–3023 (2005).
17. Krom, M. D., Emeis, K.-C. & Van Cappellen, P. Why is the Eastern Mediterranean phosphorus limited? Prog. Oceanogr. 85, 236–244 (2010).
18. Berman-Frank, I. & Rahav, E. in Life in the Mediterranean Sea: A Look at Habitat Changes 199–226 (Nova Science Publishers, 2012).
19. Man-Aharonovich, D., Kress, N., Bar-Zeev, E., Berman-Frank, I. & Béjà, O. Molecular ecology of nifH genes and transcripts in the Eastern Mediterranean Sea. Environ. Microbiol. 9, 2354–63 (2007).
20. Yogev, T. et al. Is dinitrogen fixation significant in the Levantine Basin, East Mediterranean Sea? Environ. Microbiol. 13, 854–871 (2011).
21. Ibello, V., Cantoni, C., Cozzi, S. & Civitarese, G. First basin-wide experimental results on N2 fixation in the open Mediterranean Sea. Geophys. Res. Lett. 37, 1–5 (2010).
22. Bonnet, S. et al. Dynamics of N2 fixation and fate of diazotroph-derived nitrogen in a low-nutrient, low-chlorophyll ecosystem: results from the VAHINE mesocosm experiment (New Caledonia). Biogeosciences 12, 19579–19626 (2016).
23. Raveh, O., David, N., Rilov, G. & Rahav, E. The temporal dynamics of coastal phytoplankton and bacterioplankton in the Eastern Mediterranean Sea. PLoS One 10, 1–23 (2015).
24. Spatharis, S., Skliris, N. & Meziti, A. First record of a Trichodesmium erythraeum bloom in the Mediterranean Sea. Can. J. Fish. Aquat. Sci. 69, 1444–1455 (2012).
25. Drira, Z., Chaari, D., Hamza, A. & Bel Hassen, M. Diazotrophic cyanobacteria signatures and their relationship to hydrographic conditions in the Gulf of Gabes, Tunisia. J. Mar. Biol. Assoc. United Kingdom 97, 69–80 (2017).
26. Zehr, J. P. Nitrogen fixation by marine cyanobacteria. Trends Microbiol. 19, 162–73 (2011).
27. Bergman, B., Sandh, G., Lin, S., Larsson, J. & Carpenter, E. J. Trichodesmium - a widespread marine cyanobacterium with unusual nitrogen fixation properties. FEMS Microbiol. Rev. 37, 286–302 (2013).
28. Bar-Zeev, E., Avishay, I., Bidle, K. D. & Berman-Frank, I. Programmed cell death in the marine cyanobacterium Trichodesmium mediates carbon and nitrogen export. ISME J. 7, 2340–2348 (2013).
29. Mills, M. M., Ridame, C., Davey, M., La Roche, J. & Geider, R. J. Iron and phosphorus co-limit nitrogen fixation in the eastern tropical North Atlantic. Nature 429, 292–294 (2004).
30. Sanudo-Wilhelmy, S. et al. Phosphorus limitation of nitrogen fixation by Trichodesmium in the central Atlantic Ocean. Nature411, 66–69 (2001).
31. Staal, M., Meysman, F. J. R. & Stal, L. J. Temperature excludes N2-fixing heterocystous cyanobacteria in the tropical oceans. Nature425, 504–507 (2003).
32. Capone, D. G., Zehr, J. P., Paerl, H. W., Bergman, B. & Carpenter, E. J. Trichodesmium, a globally significant marine cyanobacterium. Science. 276, 1221–1229 (1997).
33. LaRoche, J. & Breitbarth, E. Importance of the diazotrophs as a source of new nitrogen in the ocean. J. Sea Res. 53, 67–91 (2005).
34. Berman-Frank, I., Cullen, J. T., Shaked, Y., Sherrell, R. M. & Falkowski, P. G. Iron availability, cellular iron quotas, and nitrogen fixation in. Trichodesmium. Limnol. Oceanogr. 46, 1249–1260 (2001).
35. Carpenter, E. J. & Price, C. C. Nitrogen fixation, distribution and production of Oscillatoria (Trichodesmium) spp. in the western Sargasso and Caribbean seas. Limnol. Oceanogr. 22, 60–72 (1977).
36. Kress, N. & Herut, B. Spatial and seasonal evolution of dissolved oxygen and nutrients in the Southern Levantine Basin (Eastern Mediterranean Sea): chemical characterization of the water masses and inferences on the N:P ratios. Deep. Res. I 48, 2347–2372 (2001).
37. Welschmeyer, N. A. Fluorometric analysis of chlorophyll a in the presence of chlorophyll b and pheopigments. Limnol. Oceanogr.39, 1985–1992 (1994).
38. Vaulot, D. & Marie, D. Diel variability of photosynthetic picoplankton in the equatorial Pacific. Appl. Environ. Microbiol.104, 3297–3310 (1999).
39. Zehr, J. P. & McReynolds, L. A. Use of degenerate oligonucleotides for amplification of the nifH gene from the marine cyanobacterium Trichodesmium thiebautii. Appl. Environ. Microbiol. 55, 2522–2526 (1989).
40. Caporaso, J. G. et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 7, 335–336 (2010).
41. Edgar, R. C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 26, 2460–2461 (2010).
42. Heller, P., Tripp, H. J., Turk-Kubo, K. & Zehr, J. P. ARBitrator: A software pipeline for on-demand retrieval of auto-curated nifHsequences from GenBank. Bioinformatics 30, 1–8 (2014).
43. Price, M. N., Dehal, P. S. & Arkin, A. P. FastTree: computing large minimum evolution trees with profiles instead of a distance matrix. Mol. Biol. Evol. 26, 1641–1650 (2009).
44. Steemann-Nielsen, E. On the determination of the activity for measuring primary production. J Cons Int Explor Mer 18, 117–140 (1952).
45. Simon, M., Alldredge, A. & Azam, F. Bacterial carbon dynamics on marine snow. Mar. Ecol. Prog. Ser. 65, 205–211 (1990).
46. Simon, M., Alldredge, A. & Azam, F. Protein-content and protein-synthesis rates of planktonic marine-bacteria. Mar. Ecol. Prog. Ser.51, 201–213 (1989).
47. Mohr, W., Großkopf, T., Wallace, D. W. R. & Laroche, J. Methodological underestimation of oceanic nitrogen fixation rates. PLoS One 5, 1–7 (2010).
48. Weissbach, A. et al. Phytoplankton allelochemical interactions change microbial food web dynamics. Limnol. Oceanogr. 56, 899–909 (2011).
49. Heisler, J. et al. Eutrophication and harmful algal blooms: A scientific consensus. Harmful Algae 8, 3–13 (2008).
50. Villacorte, L. O. et al. Seawater reverse osmosis desalination and (harmful) algal blooms. Desalination 360, 61–80 (2015).
51. Capone, D. G. An extensive bloom of the N2-fixing cyanobacterium, Trichodesmium erythraeum, in the central Arabian Sea. Mar. Ecol. Prog. Ser. 172, 281–292 (1998).
52. Luo, Y.-W. et al. Database of diazotrophs in global ocean: abundance, biomass and nitrogen fixation rates. Earth Syst. Sci. Data 4, 47–73 (2012).
53. Spungin, D. et al. Mechanisms of Trichodesmium; bloom demise within the New Caledonia Lagoon during the VAHINE mesocosm experiment. Biogeosciences 13, 4187–4203, doi:10.5194/bg-13-4187-2016 (2016).
54. Chang, J., Chiang, K. P. & Gong, G. C. Seasonal variation and cross-shelf distribution of the nitrogen-fixing cyanobacterium, Trichodesmium, in southern East China Sea. Cont. Shelf Res. 20, 479–492 (2000).
55. González Taboada, F., González Gil, R., Höfer, J., González, S. & Anadón, R. Trichodesmium spp. population structure in the eastern North Atlantic subtropical gyre. Deep Sea Res. Part I Oceanogr. Res. Pap. 57, 65–77 (2010).
56. Rodriguez, I. B. & Ho, T.-Y. Diel nitrogen fixation pattern of Trichodesmium: the interactive control of light and Ni. Sci. Rep. 4, 4445 (2014).
57. Béjà, O. et al. Bacterial rhodopsin: evidence for a new type of phototrophy in the sea. Science. 289, 1902–1906 (2000).
58. Muñoz-Marín, M. D. C. et al. Prochlorococcus can use the Pro1404 transporter to take up glucose at nanomolar concentrations in the Atlantic Ocean. Proc. Natl. Acad. Sci. USA 110, 8597–602 (2013).
59. Bonnet, S., Grosso, O. & Moutin, T. Planktonic dinitrogen fixation along a longitudinal gradient across the Mediterranean Sea during the stratified period (BOUM cruise). Biogeosciences 8, 2257–2267 (2011).
60. Rahav, E., Ovadia, G., Paytan, A. & Herut, B. Contribution of airborne microbes to bacterial production and N2 fixation in seawater upon aerosol deposition. Geophys. Res. Lett. 43, 1–9 (2016).
61. Devassy, V. P., Bhattathiri, P. M. A. & Qasim, S. Z. Succession of organisms following Trichodesmium phenomenon. Indian J. Mar. Sci. 8, 89–93 (1979).
62. Mulholland, M. R., Bernhardt, P. W., Heil, C. A., Bronk, D. A. & Neil, J. M. O. Nitrogen fixation and release of fixed nitrogen by Trichodesmium spp. in the Gulf of Mexico. Limnol. Oceanogr. 51, 1762–1776 (2006).
63. O’Neil, J. M. The colonial cyanobacterium Trichodesmium as a physical and nutritional substrate for the harpacticoid copepod Macrosetella gracilis. J. Plankton Res. 20, 43–59 (1998).
64. Hewson, I., Govil, S. R., Capone, D. G., Carpenter, E. J. & Fuhrman, J. A. Evidence of Trichodesmium viral lysis and potential significance for biogeochemical cycling in the oligotrophic ocean. Aquat. Microb. Ecol. 36, 1–8 (2004).
65. Berman-Frank, I., Quigg, A., Finkel, Z. V., Irwin, A. J. & Haramaty, L. Nitrogen-fixation strategies and Fe requirements in cyanobacteria. Limnol. Oceanogr. 52, 2260–2269 (2007).
66. Kranz, S. A., Sültemeyer, D., Richter, K.-U. & Rost, B. Carbon acquisition by Trichodesmium: the effect of pCO2 and diurnal changes. Limnol. Oceanogr. 54, 548–559 (2009).
67. Levitan, O. et al. Combined effects of CO2 and light on the N2-fixing cyanobacterium Trichodesmium IMS101: A mechanistic view. Plant Physiol. 154, 346–356 (2010).
68. Moore, M. C. et al. Large-scale distribution of Atlantic nitrogen fixation controlled by iron availability. Nat. Geosci. 2, 867–871 (2009).
69. Huchins, D. A., Fu, F.-X., Webb, E. A., Walworth, N. & Tagliabue, A. Taxon-specific response of marine nitrogen fixers to elevated carbon dioxide concentrations. Nat. Geosci. 6, 790–795 (2013).
70. Fu, F. X. et al. Differing responses of marine N2 fixers to warming and consequences for future diazotroph community structure. Aquat. Microb. Ecol. 72, 33–46 (2014).
71. Feng, X. et al. Mixotrophic and photoheterotrophic metabolism in Cyanothece sp. ATCC 51142 under continuous light. Microbiology156, 2566–2574 (2010).
72. Montesinos, M. L., Herrero, A. & Flores, E. Amino acid transport systems required for diazotrophic growth in the cyanobacterium Anabaena sp. strain PCC 7120. Journal of Bacteriology 177, 3150–3157 (1995).
73. Benavides, M., Berthelot, H., Duhamel, S., Raimbault, P. & Bonnet, S. Dissolved organic matter uptake by Trichodesmium in the Southwest Pacific. Sci. Rep. 7, 41315 (2017).
Reconocimientos
We would like to thank Galit Ovadia for her assistance in the DNA extractions. This work was supported by grants awarded by the Ministry of National Infrastructure, Energy and Water Resources (grant number 3-11519) to E.R., by the Ministry of Environmental Protection (grant number 145–1–2) to E.R., and by the Israel Water Authority (grant number 4501-284678) to E.B-Z. and E.R.
Información sobre el autor
National Institute of Oceanography, Israel Oceanographic and Limnological Research, Haifa, 31080, Israel
Eyal Rahav
The Jacob Blaustein Institutes for Desert Research, Zuckerberg Institute for Water Research (ZIWR), Ben-Gurion University of the Negev, Sede Boqer Campus, Beer Sheva, 84990, Israel
Edo Bar-Zeev
Contribuciones
Conceived and designed the experiments: E.R. and E.B.-Z. Performed the samplings: E.R. Analyzed the data: E.R. and E.B.-Z. Contributed reagents/ materials/analysis tools: E.R. Wrote the paper: E.R. and E.B.-Z.
Intereses
The authors declare that they have no competing interests.
Correspondencia con el autor
Correspondence to Eyal Rahav or Edo Bar-Zeev.
Traducción de la publicación en https://www.nature.com/articles/s41598-017-04622-8